
2025年7月,北京林業(yè)大學(xué)張德強(qiáng)教授課題組在New Phytologist(IF=8.1)發(fā)表了題為“Phylostratigraphic analysis revealed that ancient ohnologue PtoWRKY53 innovated a vascular transcription regulatory network in Populus"的研究論文。該研究借助DAP-seq技術(shù)揭示了全基因組復(fù)制(WGD)衍生的古老PS1 ohnologs(尤其PtoWRKY53)通過新功能化創(chuàng)新了楊樹維管轉(zhuǎn)錄網(wǎng)絡(luò),為木本植物進(jìn)化發(fā)育提供了新范式。藍(lán)景科信為該研究提供了DAP-seq技術(shù)支持。

文章主要內(nèi)容
木本植物次生維管系統(tǒng)的演化是森林生態(tài)系統(tǒng)進(jìn)化歷程中的關(guān)鍵事件,其發(fā)育不僅推動(dòng)了植物向高大化方向演進(jìn),增強(qiáng)了光合效能,還拓展了植物對(duì)生態(tài)位的適應(yīng)范圍。該系統(tǒng)由木質(zhì)部、形成層和韌皮部構(gòu)成,兼具器官整合功能與物質(zhì)高效運(yùn)輸能力。其中,初生維管組織主要承擔(dān)發(fā)育中器官的水分與養(yǎng)分縱向運(yùn)輸,而次生維管組織則是植物實(shí)現(xiàn)徑向生長及木材形成的結(jié)構(gòu)基礎(chǔ)。楊樹作為木本植物研究的模式物種,其次生維管發(fā)育的分子調(diào)控機(jī)制已得到廣泛解析,但該系統(tǒng)在進(jìn)化過程中的動(dòng)態(tài)變化規(guī)律仍有待闡明。
為了確定楊屬植物內(nèi)基因的進(jìn)化起源,作者對(duì)6個(gè)高質(zhì)量測序的楊屬基因組進(jìn)行了分析,6種楊屬基因組被分為17個(gè)系統(tǒng)發(fā)育層(PS1-PS17),PS1基因占比超51%,是基因組的主要成分。基因家族聚類分析顯示,PS1的基因家族發(fā)生了顯著擴(kuò)張,表明這些古老基因的擴(kuò)張可能塑造了楊樹進(jìn)化的遺傳學(xué)基礎(chǔ)。
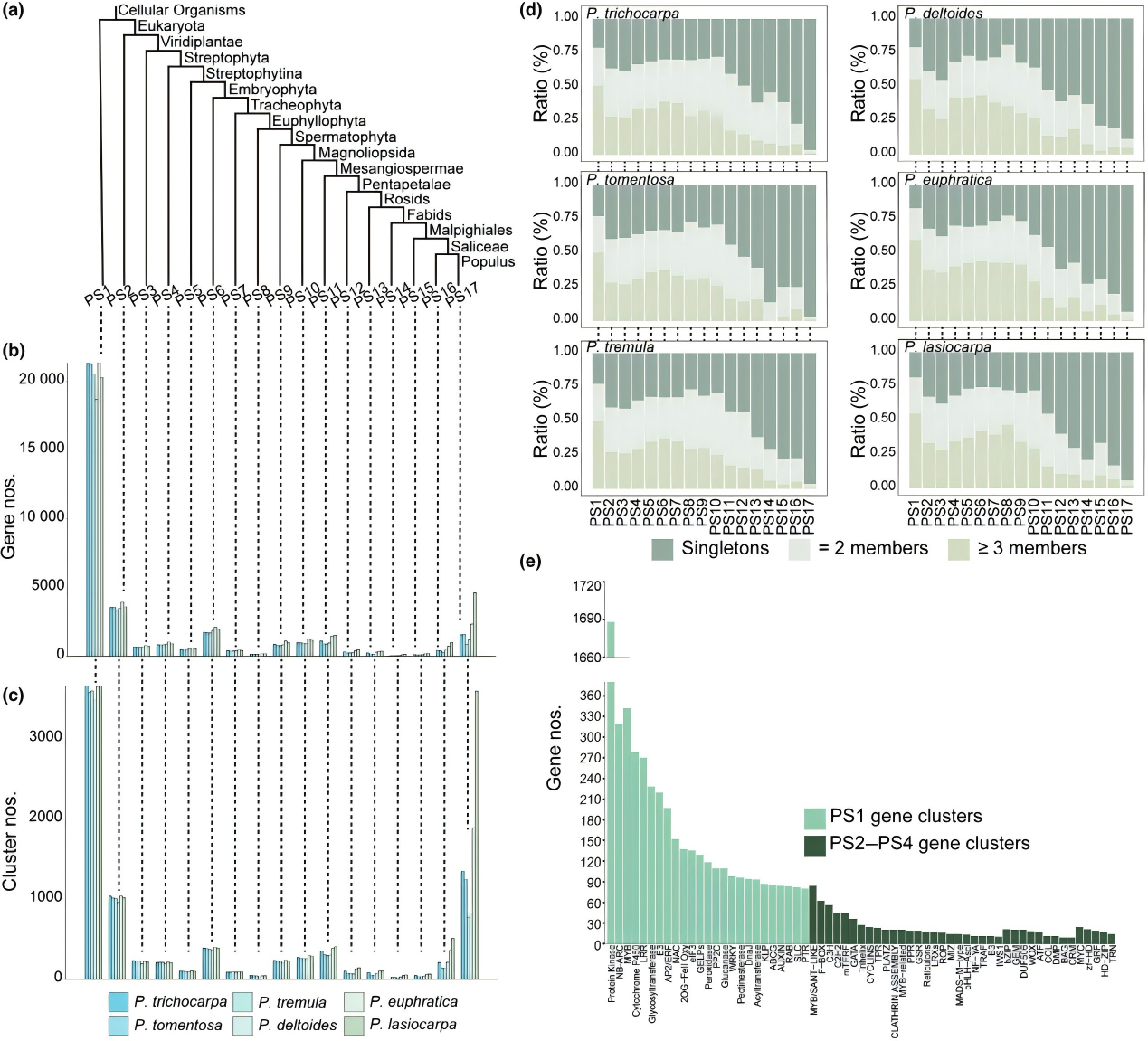
圖 1 楊屬基因組的系統(tǒng)發(fā)育特征

圖 2 不同系統(tǒng)發(fā)育層中基因的結(jié)構(gòu)及進(jìn)化特征
通過對(duì)5種楊樹維管組織(韌皮部/形成層/木質(zhì)部)進(jìn)行轉(zhuǎn)錄組學(xué)分析,并計(jì)算TAI/TDI及表達(dá)偏好指數(shù)(EP),結(jié)果顯示PS1基因占表達(dá)基因的61.55%-66.38%,且在木質(zhì)部、形成層、韌皮部中均呈現(xiàn)相似的表達(dá)水平,但轉(zhuǎn)錄分化程度差異顯著,且存在組織特異性表達(dá)模式。表明PS1基因的特異性表達(dá)驅(qū)動(dòng)了楊樹次生維管系統(tǒng)轉(zhuǎn)錄調(diào)控網(wǎng)絡(luò)的差異形成。

圖3 楊樹莖維管束組織中表達(dá)基因的進(jìn)化轉(zhuǎn)錄組圖譜
楊樹次生維管系統(tǒng)轉(zhuǎn)錄組與共表達(dá)網(wǎng)絡(luò)(WGCNA)分析顯示,劑量非限制性的PS1 ohnologues(MYB、NAC、WRKY 家族)在木質(zhì)部存在特異性表達(dá)。通過對(duì)轉(zhuǎn)基因材料進(jìn)行表型分析,結(jié)果顯示PtoWRKY53-OE 株系高度降低,莖直徑減少,木質(zhì)部寬度減少;RNAi 株系則相反。電鏡觀察表明,PtoWRKY53-OE 株系纖維細(xì)胞壁變薄,纖維素含量降低;RNAi 株系則相反,證實(shí)PtoWRKY53抑制木質(zhì)部次生壁沉積。
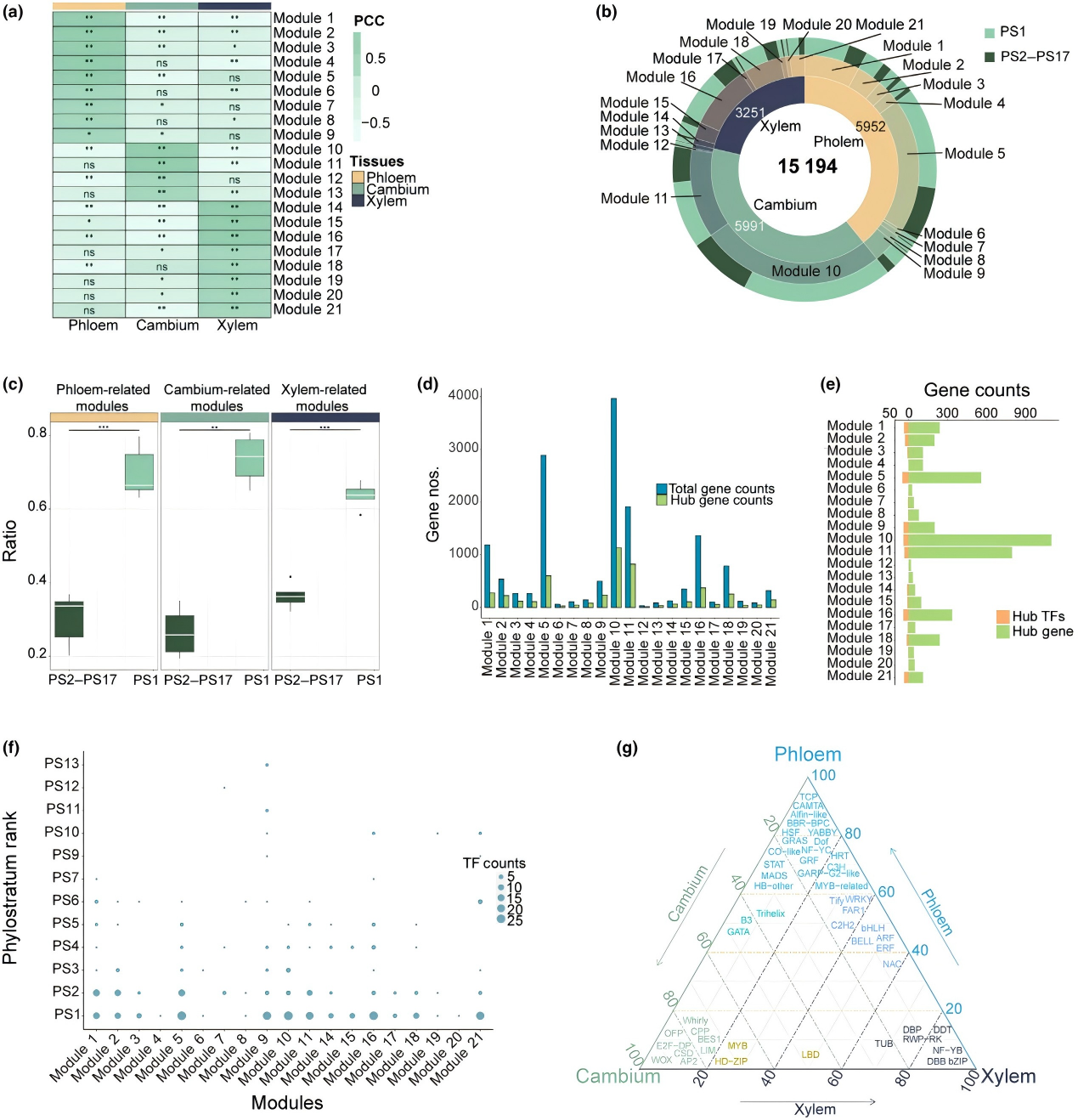
圖 4 楊屬植物維管組織的共表達(dá)網(wǎng)絡(luò)分析
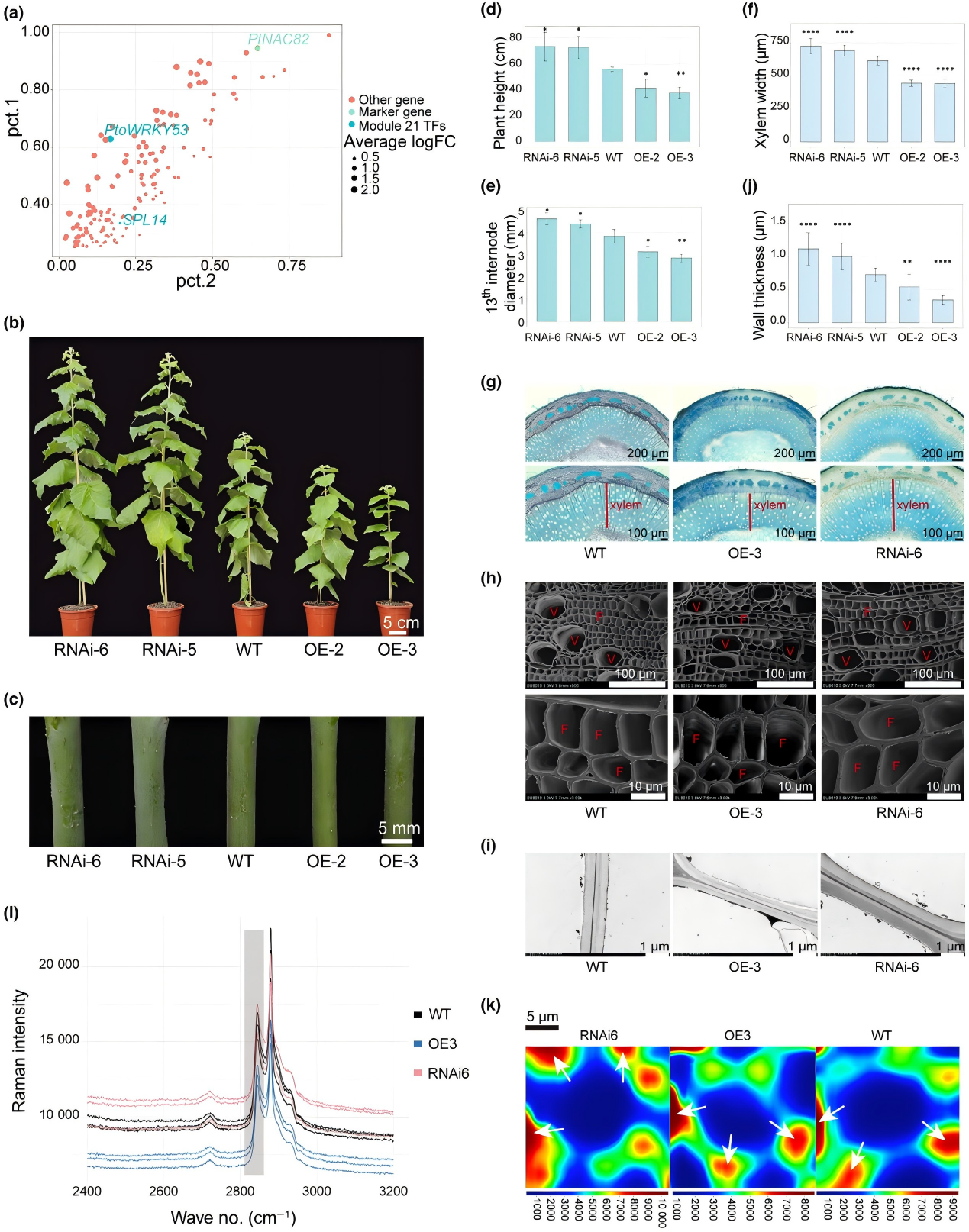
圖 5 PtoWRKY53 對(duì)木質(zhì)部細(xì)胞次生細(xì)胞壁發(fā)育的影響
進(jìn)一步通過DAP-seq探究PtoWRKY53 在木質(zhì)部發(fā)育中的直接靶標(biāo)。發(fā)現(xiàn)在下游靶基因的啟動(dòng)子區(qū)域存在464個(gè)共同結(jié)合位點(diǎn)。順式作用元件W-box(TTGAC)始終位于每個(gè)峰的中心,且在兩個(gè)重復(fù)實(shí)驗(yàn)中表現(xiàn)出很強(qiáng)的可重復(fù)性。差異表達(dá)分析結(jié)果表明,Potri.016G054900(PtrCESA3)、Potri.004G118700(PtrTPXL5)、Potri.002G254800和Potri.006G047300直接受其調(diào)控。已知CESA3在擬南芥中影響纖維素合成,在楊樹中可能發(fā)揮類似作用。這表明PtoWRKY5可能直接調(diào)控PtoCESA3的表達(dá)。后續(xù)通過雙熒光素酶報(bào)告實(shí)驗(yàn)、EMSA和Y1H證實(shí),PtoWRKY53通過結(jié)合PtoCESA3啟動(dòng)子的W-box抑制其轉(zhuǎn)錄,減少了木質(zhì)部纖維細(xì)胞次生壁的沉積,最終負(fù)調(diào)控楊樹木質(zhì)部發(fā)育。
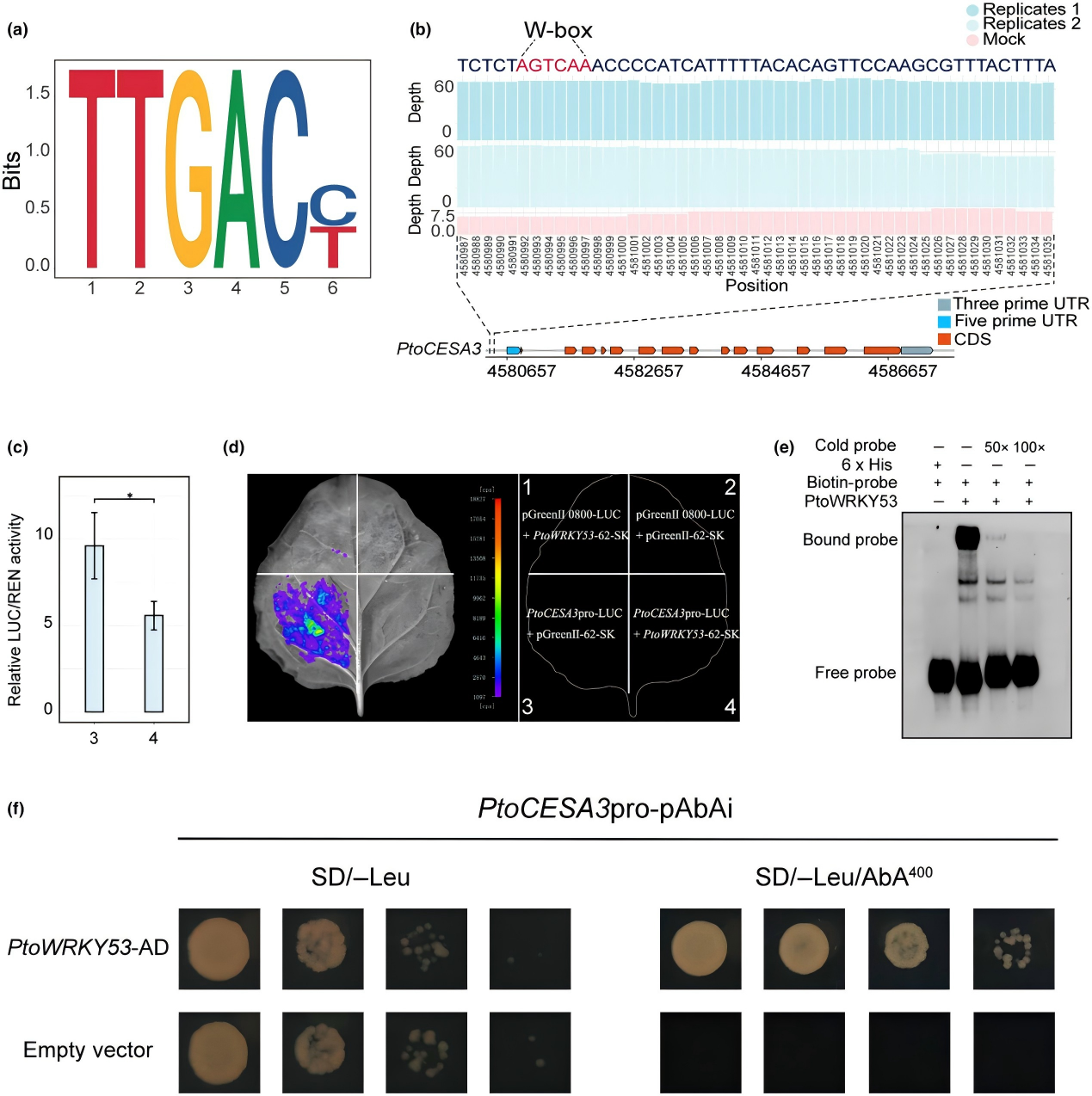
圖 6 PtoWRKY53 負(fù)調(diào)控 PtoCESA3
本研究發(fā)現(xiàn),5種楊樹中,35個(gè)基因受PtoWRKY53保守調(diào)控,其中29個(gè)為PS1基因,包括直接靶基因 PtoCESA3、PtoTPXL5等。啟動(dòng)子分析顯示,PtoCESA3的W-box在楊屬中保守(除胡楊),在擬南芥中缺失,表明PtoWRKY53-PtoCESA3模塊是楊屬的新調(diào)控機(jī)制。
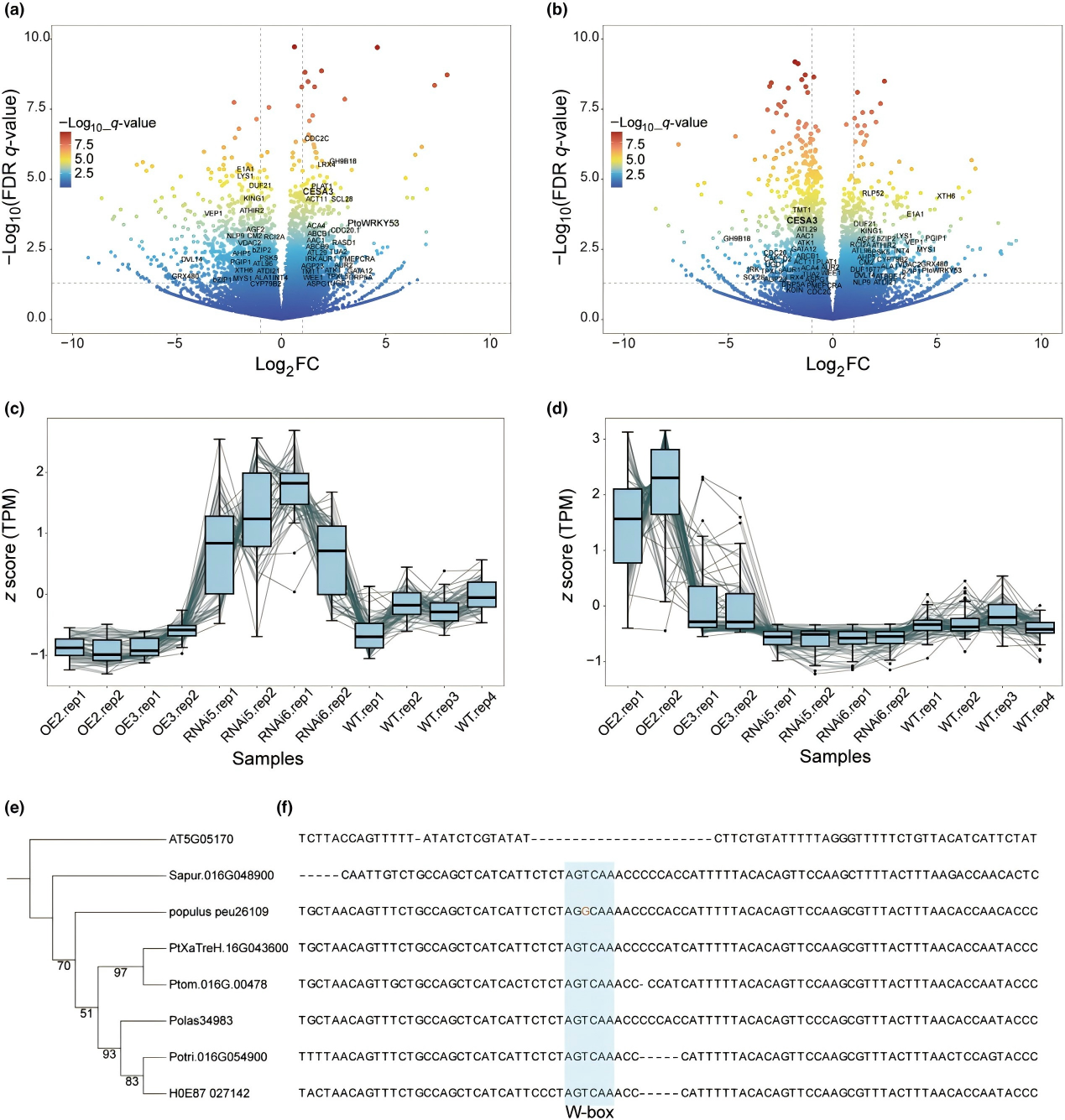
圖 7 PtoWRKY53 在楊屬植物中形成保守的轉(zhuǎn)錄調(diào)控網(wǎng)絡(luò)
該研究揭示了楊樹次生維管發(fā)育調(diào)控機(jī)制地點(diǎn)演化路徑,系統(tǒng)構(gòu)建了楊樹維管系統(tǒng)發(fā)育的遺傳調(diào)控網(wǎng)絡(luò),深入解析了PtoWRKY53在次生木質(zhì)部發(fā)育中的功能機(jī)制,為深入理解多年生樹木次生維管系統(tǒng)的進(jìn)化奠定了重要理論基礎(chǔ)。
發(fā)布DAP文章最后附圖.jpg")
