
2025年9月27日,中國農(nóng)業(yè)大學(xué)孫其信院士/邢界文教授團(tuán)隊在Molecular Plant(IF: 24.1)在線發(fā)表了題為An incoherent feed-forward loop coordinates nitrate uptake and tillering in wheat的研究論文,該研究揭示了小麥中TaNLP3通過非一致性前饋環(huán)(incoherent feed-forward loop)協(xié)同調(diào)控硝酸鹽吸收與分蘗的分子機(jī)制,還鑒定出可直接用于育種的優(yōu)異單倍型。藍(lán)景科信為該研究提供了DAP-seq技術(shù)支持。

文章主要內(nèi)容

小麥作為近20%人口的主食來源,其產(chǎn)量和氮素利用效率(NUE)直接關(guān)系到糧食安全與農(nóng)業(yè)可持續(xù)發(fā)展。自綠色革命以來,氮肥施用大幅提升了小麥產(chǎn)量,但如今增產(chǎn)效應(yīng)已進(jìn)入平臺期——過量施氮不僅降低NUE,還會催生大量無效分蘗,造成資源浪費。如何讓小麥在氮素波動環(huán)境中“智能"平衡硝酸鹽吸收與有效分蘗,成為小麥遺傳改良中的核心問題。
主要研究結(jié)果
核心發(fā)現(xiàn)一:關(guān)鍵基因鑒定與功能解析——鎖定硝酸鹽響應(yīng)核心因子
1、關(guān)鍵標(biāo)記基因鑒定:TaNRT2.1-6B4 是氮響應(yīng)與分蘗的“信號樞紐"
本研究通過對缺氮后復(fù)氮處理的小麥幼苗進(jìn)行不同時間點(15min、1h、6h)RNA-seq分析,結(jié)合GO富集分析與候選基因關(guān)聯(lián)分析,鑒定到TaNRT2.1-6B4為小麥硝酸鹽響應(yīng)關(guān)鍵標(biāo)記基因,其表達(dá)量與分蘗數(shù)呈顯著正相關(guān),且受硝酸鹽強(qiáng)烈誘導(dǎo)(轉(zhuǎn)錄水平提升5-180倍),成為連接氮吸收與分蘗發(fā)育的關(guān)鍵“信號樞紐"。
2、核心調(diào)控因子鑒定:TaNLP3 是平衡氮吸收與分蘗的“總開關(guān)"
通過酵母單雜篩選TaNRT2.1-6B4的上游調(diào)控因子,團(tuán)隊鑒定出NIN-like蛋白家族成員TaNLP3。轉(zhuǎn)錄組數(shù)據(jù)顯示,TaNLP3不受短期施氮誘導(dǎo),且在不同氮磷供給組合條件下表達(dá)量保持穩(wěn)定。亞細(xì)胞定位、瞬時熒光素酶(LUC)、酵母轉(zhuǎn)錄激活實驗等技術(shù)驗證基因表達(dá)特征與轉(zhuǎn)錄激活功能。結(jié)果顯示TaNLP3作為硝酸鹽信號通路的核心轉(zhuǎn)錄因子,定位于細(xì)胞質(zhì)與細(xì)胞核,硝酸鹽處理可促進(jìn)其核滯留,其C端具有轉(zhuǎn)錄激活活性,N端片段次之,全長蛋白的激活活性最弱。

圖1 TaNRT2.1-6B4 是受TaNLP3正調(diào)控的關(guān)鍵硝酸鹽響應(yīng)標(biāo)記基因
利用CRISPR-Cas9技術(shù)構(gòu)建TaNLP3敲除株系,通過田間試驗(正常氮/低氮條件)與水培試驗,系統(tǒng)分析植株表型、氮含量及15N吸收速率,結(jié)果顯示TaNLP3敲除顯著降低小麥株高、分蘗數(shù)及產(chǎn)量,且在正常氮條件下表型缺陷更顯著,但植株氮含量未受影響;此外在硝酸鹽誘導(dǎo)(NI)條件下敲除TaNLP3抑制硝酸鹽吸收,而在低氮(LN)和正常氮(NN)穩(wěn)態(tài)條件下不影響甚至促進(jìn)吸收。表型分析表明,TaNLP3以氮素依賴的方式調(diào)控硝酸鹽吸收和小麥分蘗——在硝酸鹽誘導(dǎo)(短期氮信號)下,它主要促進(jìn)硝酸鹽吸收;在正常氮供應(yīng)(長期氮信號)下,它更側(cè)重調(diào)控分蘗發(fā)育,適配小麥在不同氮環(huán)境中的生長需求。

圖2 TaNLP3在短期和長期硝酸鹽處理下調(diào)控不同基因的表達(dá)
核心發(fā)現(xiàn)二:硝酸鹽信號調(diào)控網(wǎng)絡(luò)構(gòu)建——解析協(xié)同調(diào)控分子路徑
1、TaNLP3–TaLBD38–TaNRT2.1 構(gòu)成非一致性前饋環(huán)路
為解析TaNLP3在短期和長期硝酸鹽信號中功能背后的調(diào)控網(wǎng)絡(luò),作者對TaNLP3敲除株系與Fielder對照在不同氮條件(NN、NS、NI)下的根系和地上部分進(jìn)行轉(zhuǎn)錄組測序(RNA-seq),鑒定到調(diào)控小麥分蘗過程中的關(guān)鍵下游候選基因TaLBD38,原位雜交結(jié)果顯示,TaLBD38與TaNLP3均在小麥分蘗芽中表達(dá),隨后通過對正常氮條件下生長的Fielder 和Tanlp3-4的分蘗芽進(jìn)行了轉(zhuǎn)錄組測序驗證了TaNLP3–TaLBD38調(diào)控級聯(lián)在分蘗發(fā)育中的作用。
不同氮素條件下TaNRT2.1家族基因與TaLBD38的表達(dá)熱圖顯示,二者具有基因特異性的硝酸鹽響應(yīng)模式,結(jié)合此前研究證實LBD家族成員參與氮素利用和地上部分枝調(diào)控,作者提出:小麥硝酸鹽信號通路中,TaNLP3、TaLBD38與TaNRT2.1構(gòu)成非一致性前饋環(huán)路:短期信號中,TaNLP3感知硝酸鹽信號后進(jìn)入細(xì)胞核,激活TaNRT2.1以增強(qiáng)硝酸鹽吸收,同時誘導(dǎo)TaLBD38 表達(dá);長期信號傳導(dǎo)中,積累的TaLBD38蛋白會抑制TaNRT2.1的表達(dá),避免硝酸鹽過量吸收,進(jìn)而促進(jìn)分蘗形成。進(jìn)一步EMSA、LUC、ChIP-qPCR等分子實驗表明TaNLP3可結(jié)合并激活TaNRT2.1-6B4和TaLBD38-4A的啟動子;而TaLBD38則結(jié)合并抑制TaNRT2.1-6B4的啟動子。EMS突變體的RT-qPCR、表型與氮含量測定證實TaLBD38缺失會致氮吸收異常與發(fā)育缺陷。上述結(jié)果均證明該環(huán)路實現(xiàn)了硝態(tài)氮吸收與分蘗形成的動態(tài)平衡。

圖3 TaNLP3通過非一致性前饋環(huán)調(diào)控TaNRT2.1與TaLBD38的表達(dá),進(jìn)而協(xié)同調(diào)控硝酸鹽吸收與分蘗
2、TaLBD38通過抑制TaCKX4/5調(diào)控分蘗
為篩選TaLBD38介導(dǎo)的分蘗調(diào)控關(guān)鍵基因,作者對野生型和3個TaLBD38 RNAi株系10天齡幼苗進(jìn)行了轉(zhuǎn)錄組測序。根的轉(zhuǎn)錄組分析結(jié)果顯示,TaLBD38調(diào)控的差異基因廣泛參與養(yǎng)分利用過程,包括硝酸鹽轉(zhuǎn)運與同化、氨基酸轉(zhuǎn)運、鐵離子轉(zhuǎn)運及磷酸鹽調(diào)控。與根系DEGs不同,莖部DEGs富集于BR、赤霉素(GA)和細(xì)胞分裂素(CK)信號通路。由于TaLBD38是一種轉(zhuǎn)錄抑制因子,其直接調(diào)控靶點在RNAi細(xì)胞系中被上調(diào)。根據(jù)這一標(biāo)準(zhǔn),結(jié)合水稻相關(guān)研究推斷出TaCKX4/5是介導(dǎo)TaLBD38調(diào)控分蘗的主要下游靶點。隨后通過RT-qPCR,LUC和EMSA實驗證實TaLBD38直接結(jié)合并抑制細(xì)胞分裂素降解酶基因TaCKX4/5的啟動子,提高局部細(xì)胞分裂素水平,促進(jìn)分蘗形成。

圖4 TaLBD38通過在根部抑制TaNRT2.1家族基因的表達(dá)以限制硝酸鹽吸收,同時在地上部抑制TaCKX4/5的表達(dá)以促進(jìn)分蘗
核心發(fā)現(xiàn)三:染色質(zhì)層面調(diào)控——TaNLP3與SWI/SNF復(fù)合物協(xié)同“解鎖"靶基因
為了揭示TaNLP3調(diào)控基因表達(dá)的深層機(jī)制,作者以TaNLP3為誘餌進(jìn)行Y2H實驗,鑒定出62個候選相互作用物。其中,SWI/SNF染色質(zhì)重塑復(fù)合物為關(guān)鍵物質(zhì)。Co-IP、LUC和雙分子熒光互補(bǔ)實驗進(jìn)一步證實TaNLP3無論體內(nèi)體外均可與SWI/SNF亞基TaSYD、TaSNF5直接相互作用。

圖5 SWI/SNF 染色質(zhì)重塑復(fù)合體與 TaNLP3 相互作用,調(diào)控小麥硝酸鹽響應(yīng)
為了研究TaNLP3敲除是否在全基因組水平上影響染色質(zhì)狀態(tài),團(tuán)隊運用轉(zhuǎn)座酶可及性染色質(zhì)測序(ATAC-seq)分析TaNLP3敲除株系與Fielder對照在NS和NI條件下的全基因組染色質(zhì)可及性差異。結(jié)果顯示TaNLP3是硝酸鹽誘導(dǎo)的染色質(zhì)重塑關(guān)鍵調(diào)控因子,并且鑒定到23365個受硝酸鹽和TaNLP3共同調(diào)控的可及染色質(zhì)區(qū)域。為進(jìn)一步明確哪些差異可及染色質(zhì)區(qū)域可能受TaNLP3直接調(diào)控,作者進(jìn)行了DNA親和純化測序(DAP-seq)鑒定TaNLP3的直接靶基因。兩個重復(fù)交集中鑒定到18941個靶基因,這些18941個基因共鑒定出54018個峰,其中90%的結(jié)合峰位于轉(zhuǎn)錄起始位點(TSS)上游5kb調(diào)控區(qū)。整合ATAC-seq和DAP-seq數(shù)據(jù)分析發(fā)現(xiàn),4008個基因既與硝酸鹽和TaNLP3共同調(diào)控的可及染色質(zhì)區(qū)域相關(guān),又是DAP-seq鑒定的TaNLP3直接靶基因,這些重疊基因中包含許多氮素相關(guān)基因,包括參與硝酸鹽吸收、硝酸鹽同化和硝酸鹽信號調(diào)控的基因。IGV峰圖,顯示TaNRT2.1-6B4和TaLBD38-4A位點的染色質(zhì)可及性顯著提高,且存在TaNLP3強(qiáng)結(jié)合信號,這些位點的DAP-seq峰為TaNLP3直接結(jié)合并調(diào)控這些關(guān)鍵硝酸鹽響應(yīng)基因的啟動子提供了有力證據(jù)。染色質(zhì)可及性的增加通常伴隨著活躍的表觀遺傳修飾,因此檢測了四個氮相關(guān)基因(TaNRT2.1-6B4、TaNIA1-6A、TaNIA1-6B 和TaLBD38-4A)啟動子區(qū)域的H3K9ac水平。ChIP–qPCR結(jié)果顯示NI條件下,F(xiàn)ielder中H3K9ac水平顯著高于NS條件;而敲除TaNLP3后,NI條件下H3K9ac水平大幅下降。

圖6 TaNLP3 通過調(diào)控其靶基因處的染色質(zhì)可及性,進(jìn)而調(diào)控硝酸鹽響應(yīng)基因的表達(dá)
核心發(fā)現(xiàn)四:育種新資源——3個優(yōu)異單倍型實現(xiàn)“高產(chǎn)+高NUE"
為評估TaNLP3-TaLBD38-TaNRT2.1調(diào)控模塊的育種潛力,團(tuán)隊對405份小麥種質(zhì)資源(359個栽培種、46個地方品種)進(jìn)行單倍型分析,鑒定出3個關(guān)鍵基因的優(yōu)異單倍型:TaNLP3-3BHap2、TaLBD38-4AHap2 和 TaNRT2.1-6B4Hap1。這些優(yōu)異單倍型在現(xiàn)代栽培種中的頻率呈下降趨勢(部分從70%降至30%),這為通過分子標(biāo)記輔助育種、回補(bǔ)優(yōu)異單倍型提供了重要靶點——將這些單倍型聚合到同一品種中,有望培育出“低氮高產(chǎn)、高氮高效"的小麥新品種。

圖7 小麥種質(zhì)資源中TaNLP3、TaLBD38 及 TaNRT2.1的單倍型分析
小結(jié)
該研究闡明了TaNLP3作為核心調(diào)控因子,通過與SWI/SNF復(fù)合體協(xié)作重塑染色質(zhì)可及性,借助不一致性前饋環(huán)實現(xiàn)硝酸鹽吸收與分蘗形成的時空協(xié)同。不僅深化了對作物氮信號網(wǎng)絡(luò)的理解,也為通過分子設(shè)計培育“高產(chǎn)高效"小麥品種提供了新靶點與遺傳資源。
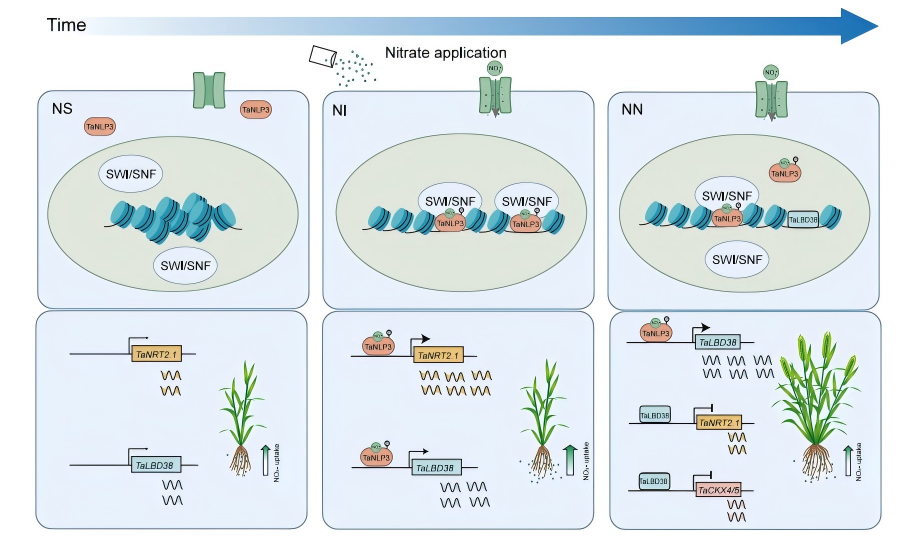
圖8 TaNLP3介導(dǎo)的染色質(zhì)可及性調(diào)控及小麥氮-分蘗平衡模型
布DAP文章最后附圖.jpg")
